
専門家向け研究紹介(概要)
ミトコンドリア形態制御の分子機構と生理機能
ミトコンドリアの融合と分裂を制御するGTP加水分解酵素(GTPase)
ミトコンドリアはダイナミックなオルガネラであり、融合が活性化するとより長くつながった構造が、逆に分裂が活性化すると小さな断片化したミトコンドリアが形成される。近年、局在の異なるGTPase群がミトコンドリア融合・分裂に関与することがわかってきた(図1)。私たちはこれらのGTPaseの解析を中心として、ミトコンドリア膜ダイナミクスの分子機構解明を目指して研究を行っている。

図1 ミトコンドリア形態制御に機能するGTPaseたち
Drp1は細胞質・外膜に局在し、分裂に機能する。Mfn1/Mfnは外膜に局在し融合に機能する。
OPAは膜間スペースに局在し、融合及び内膜クリステ構造形成に機能する。
哺乳動物ミトコンドリアの融合反応の可視化
ミトコンドリアは外膜と内膜の2つの膜から形成されており、ミトコンドリア融合時には外膜融合に引き続いて内膜の融合がおこる。私達はミトコンドリアを蛍光蛋白質で標識し生細胞観察を行うことにより、ミトコンドリアは頻繁に融合し、その内容物を交換できることを明らかにしている(図2)。

図2 ミトコンドリア融合反応の可視化
ミトコンドリアを異なる蛍光蛋白質で可視化して細胞融合する(融合前:左)
生細胞観察を行うと2色のミトコンドリアが融合していく過程を観察できる(中央から右へ)
Mitofusin (Mfn)によるミトコンドリアの融合
ミトコンドリア外膜の融合には外膜のGTPase、 Mitofusin (Mfn)が機能する。私達は哺乳動物 から2つのMfnアイソフォーム(Mfn1, Mfn2)を単離・解析し、2つのMfnアイソフォームは機能分担しており、協調的に機能してミトコンドリア融合を制御していることを示している。
さらにMfn蛋白質の機能を生化学的に解析を行ったところ、Mfn1はGTP加水分解に依存して2つのミトコンドリアを結合させる機能を持っていることが明らかになった(図3)。一方構造解析から、Mfn1のC末端領域は細胞質で二量体を形成することも報告されている。しかし、この後の膜融合の分子機構、Mfn2の機能など、ミトコンドリア融合反応には明らかにすべき重要な問題が多く残されている。

図3 Mfn1によるミトコンドリアの結合
GTPの加水分解に依存してミトコンドリアを結合する
OPA1の切断とミトコンドリア融合制御
ミトコンドリア膜間スペースに局在するダイナミン様GTPase、OPA1は神経形成異常となる神経変性疾患 Dominant Optic Atrophy type Iから同定された、ミトコンドリア融合に関与する蛋白質である。
上記図1のミトコンドリア融合解析の過程で、私たちはミトコンドリア内膜の膜電位がミトコンドリア融合に必須であることに気が付いた。膜電位によるミトコンドリア融合の制御機構を解析した結果、膜電位が消失するとOPA1がミトコンドリア内で切断され、その活性を失うことを見出した(図4)。この切断はアポトシス誘導時にも観察される。つまりOPA1はミトコンドリア呼吸活性のセンサーとして機能し、活性が落ちたミトコンドリアの融合を 押さえていると考えられる。「働きの悪いミトコンドリアは細胞内のミトコンドリアネットワークから排除してしまう」 ことになるため、ミトコンドリアの品質管理にとって都合の良い機構であるといえるだろう。

図4 OPA1の切断によるミトコンドリア形態制御
膜電位の消失に伴いOPA1が切断され、ミトコンドリア融合が抑制される。
ミトコンドリアの分裂、細胞分裂期のミトコンドリア形態変化
ミトコンドリアは細菌(α-プロテオバクテリア)の共生を起源にしたオルガネラであると考えられている。共生の初期段階には細菌型の分裂を行っていたようだが、酵母や哺乳動物では細菌型の分裂装置は見られず共生後に新たな分裂システムを獲得した。細胞質に局在するダイナミン様GTPase・Drp1とミトコンドリア外膜蛋白質Fis1がミトコンドリア分裂に機能することが明らかになっている(図1)。
細胞分裂期のミトコンドリアのダイナミクスを観察したところ、細胞分裂期にはミトコンドリアが小さく断片化して、娘細胞に分配されたあとで長いミトコンドリアのネットワーク構造が再び形成されることに気が付いた(図5)。このミトコンドリア分裂はDrp1の機能に依存していた。
さらに私達は、細胞分裂期にはDrp1がMPF (Cdk1/cyclinB)によって1箇所だけ特異的にリン酸化され、活性化することによって、ミトコンドリアが分裂することを明らかにしている。この発見の後に、Drp1はリン酸化のみならずユビキチン化、SUMO化などさまざまな翻訳後修飾を受けて機能が制御されていることが明らかにされてきている。

図5 細胞分裂期のミトコンドリア分裂
細胞分裂期にはミトコンドリア分裂因子Drp1がリン酸化され、
分裂が促進されるためミトコンドリアが小さく断片化する。
一方、Drp1発現抑制細胞では細胞分裂期でも長いミトコンドリアが観察される。
ミトコンドリアDNAの「核様体構造」の形成とそのダイナミクス
ヒトでは、細胞あたり数百コピー以上の環状のmtDNAを保持している。私達はmtDNAのライブイメージング系を構築しており、ミトコンドリアの膜とDNAは協調的に制御されていること、mtDNAの配置が心筋の成長など個体レベルでも重要な役割を持つことなどを明らかにしている。このmtDNAの個体内での遺伝様式を知ることは、病気や老化におけるミトコンドリアの役割を知るうえで重要な意味を持つのではないかと考えて研究を進めている。
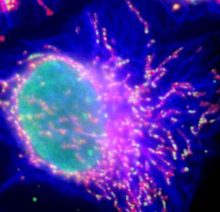
図6 ミトコンドリア内のDNAの局在。
哺乳動物培養細胞のミトコンドリア(赤)とDNA(緑)を同時に染色して蛍光顕微鏡で観察すると、
ミトコンドリア上にドット状に見える多くの構造が観察される。これは核様体と呼ばれているが、
その形成の分子機構や変動の意義はよくわかっていない。なお同時に微小管を青色で示している。
無断転載を禁じます